|
||||
A Biologically Inspired Mechano-optical Imaging System Based in Insect Vision |
Susanne K. Williams BSc, BAppSci (Photography) David Reser Ph.D and Adrian G. Dyer BAppSci (Photography), Ph.D Nature has devised a number of imaging systems that allow animals to view their surroundings. Both the lens eye, as is found in most vertebrates, and the pinhole eye found in Nautilus, have been used as models for photographically reproducing visual information. Most invertebrates (e.g. bees and flies) image their world with compound eyes, and until recently the compound eye design has been largely ignored as an option for photographic imaging. In this study we investigated the principle of making photographs with a mechano-optical device that accurately simulates the frontal foveal region of a bee vision where ommatidia (facets) are approximately parallel. For low spatial frequency information (< 0.2 cycles/degree) the system reliably resolves subject detail, but there is a rapid decrease in the transfer of spatial information at higher frequencies because (1) the quanta of photons from respective parts of a scene become indistinguishable from the random noise inherent in the system, and (2) a point in a scene becomes progressively imaged by an increasing number of ray selectors as its distance from the mechano-optical device increases. The findings are consistent with Shannon’s sampling theory and suggest that this type of imaging device operates best for small working distances between the subject and optical device. Because of the sharp decrease in the modulation of image contrast with increasing spatial frequencies the optical device also acts as an effective low pass filter which freely transmits all wavelengths of radiation.
|
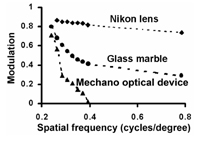
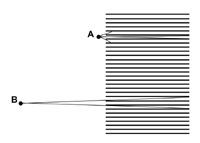
Introduction The recent, rapid advances in digital imaging technology have lead to the miniaturisation of optics and sensors for a wide range of applications like mobile phones (Tanida et al. 2003). In nature, animals have evolved a number of optical solutions for imaging their surroundings and these have been useful models for the development of modern photographic optics (Wald 1950). For example, animals like the mollusc Nautilus use pinhole optics to view the world (Muntz and Raj 1984), while most vertebrates use a simple lens to focus an image onto a retina. Both of these principles have important applications in imaging science (Wald 1950, Stroebel et al. 1996, Renner 1999). Invertebrates, including bees and flies, view their world using a different optical principle in which different parts of a scene are viewed by individual units called ommatidia. Each ommatidium has only a narrow range of angular acceptance, but integrating how the world is sampled by a large number of ommatidia permits a coherent representation of a scene (Dyer and Williams 2005). The number of ommatidia per eye is proportional to the square root of both body size and eye size of individual bees (Jander and Jander 2002). For example, each apposition compound eye of the honeybee contains approximately 4500 ommatidia, which each selectively transmit information from one small part of a scene; but the integration of all of the individual images allows the bee to spatially resolve visual information about the surrounding world (Land 1997). One recent study has been able to fabricate artificial polymer ommatidia that have potential applications in medical, industrial and military areas (Jeong et al. 2006). To understand how this type of optical principle might be useful as a photographic imaging tool, it is important to understand how a mechano-optical device modulates image contrast, and how different components of a scene might be processed in animals like bees. For example, a recent study that investigated how visual scenes might be viewed considering the frontal foveal region of the honeybee eye, where ommatidia are approximately parallel, concluded that rather than viewing a faceted image, bees probably view a coherent image because the bee brain would most likely fill in components of an image between adjacent ommatidia (Dyer and Williams 2005). However, that study also illustrated that the modulation of image contrast for square wave gratings rapidly declined when compared with simple lens optics represented by either a glass marble or an engineered photographic lens. Objectives This study used a mechano-optical principle (Knowles and Dartnall 1977, Williams and Dyer 2002, Dyer and Williams 2005, Williams and Dyer 2007) to image a representation of a flower at a number of different distances. This was to establish how image quality with a mechano-optical device is affected by viewing distance. The study then considered the main optical factors that influence image quality for this type of optical device. Methodology Mechano-optical device An optical device to simulate how the apposition compound eye of honeybees resolves visual information was constructed using an array of parallel mounted straws. A total of 4500 black drinking straws of 240 mm length and 4 mm diameter with angular resolution of 0.95 degrees of visual arc; (Dyer and Williams 2005) were stacked into a 36 x 38 cm wooden frame to simulate the frontal foveal region of an insect’s eye, where the interommatidial angle is relatively small and ommatidia are approximately parallel to each other (Dyer and Williams 2005). Using a collection of straws of this dimension enabled each optical waveguide for the mechano-optical system to accurately represent the resolution of the eye of the honeybee. The resolution of the system for resolving square wave gratings has previously been measured (Figure 1) and closely matches behavioural data for honeybees discriminating square wave grating presented in a Y-maze apparatus (Srinivasan and Lehrer 1988). To photographically record images formed by the 4500, straws a sheet of architectural drafting paper was mounted directly behind the straws so that an object placed in front of the straw camera formed an image on the drafting paper. The image projected onto the drafting paper was then subsequently re-photographed using a Nikon 995 (3.34 megapixel) digital camera (Nikon, Japan) as an uncompressed grey scale TIFF file. Image processing was subsequently done on a desktop computer using Adobe™ Photoshop™ software (Adobe Systems Incorporated, North America). Stimuli recording An achromatic photograph of a cape dandelion (Arctotheca calendula) flower was printed on matt photographic paper at a size of 35 cm diameter between opposing petals. The resulting print was then illuminated with copy lighting and re-photographed through the mechano-optical device at distances of 10, 20, 30, 40, 50, and 60 cm from the front edge of the mechano-optical device. For each of these distances, the approximate spatial frequency (considering the seven cycles in petal frequency from the top to the bottom of the image for each distance) are respectively; 0.06, 0.08, 0.12, 0.15, 0.18, 0.22. Thus, by photographing the flower image at different distances from the optical device, it is possible to illustrate how the image contrast modulation for a natural scene image, such as a flower, varies depending upon subject distance from the optical device. Ray tracing A ray tracing diagram was used to demonstrate the optical principles of the mechano-optical device as it clearly illustrates the sharp drop in modulation of image contrast (as compared with a lens system). Two potential problems were considered. First, how the quanta of photons from respective parts of a scene become indistinguishable from the random noise inherent in the system as the subject matter being imaged becomes smaller. Second, how a point in a scene becomes progressively imaged by an increasing number of ray selectors as the viewing distance from the mechano-optical device increases. Results and Discussion Figure 2 A-F shows images of the flower stimulus taken through the mechano-optical array at a variety of different distances. At the closest distance flower shape is clear- including detail of the stamen, but at the furthest distance it is only just possible to resolve some of the inner flower structure from the outer flower structure (individual petals are no longer resolved). This decline in the spatial representation of the flower occurs over a relatively short (compared with a lens) working distance (10-60 cm), which can also be represented by the steep decline in the modulation transfer function (mtf) of the mechano-optical device compared with a simple lens (Figure 1). The upper limit on spatial frequency that can be resolved using the mechano-optical array is determined by the length and diameter of each array element. In this case, the length was selected such that each ‘ommatidium’ has an angular resolution of 0.95 degrees of visual arc. The sampling theorem of Shannon (1998) predicts that the theoretical cut off is when half the information from the scene is successfully transmitted, yielding a limit of approximately 0.47 cycles per degree (cpd) (Dyer and Williams, 2005). Empirical measurements of the mechano optical array for square wave gratings agree well with this theoretical limit (Dyer and Williams, 2005). In this current study Figures 2 E-F indicate that for natural stimuli like flowers the maximum resolved spatial frequency is about 0.22 cpd. The variations in this result compared with those of Dyer and Williams (2005) are probably due to differences in contrast for either a black and white square wave grating or the comparatively low contrast natural image of a flower. As spatial frequency increases linearly with distance from the viewer, performance is rapidly degraded as the device is moved away from the stimulus, and interestingly this sharp decrease in the modulation of image contrast with increasing spatial frequencies potentially allows the mechano-optical device to act as an effective low pass filter that freely transmits all wavelengths of radiation (since there is actually no medium, other than air or even potentially a vacuum, that radiation must pass through). The model we examined in the current study investigates the optical properties of a mechano-optical device considering only achromatic light. There is no attempt to model bee colour vision in this study, as the colour simulation of bee vision has recently been demonstrated photographically (Williams and Dyer 2007). There are two main factors that contribute to the sharp decline in the mtf of the mechano-optical array. The first factor is that the quanta of photons from respective parts of a scene become indistinguishable from the random noise inherent in the system (Figures 2 G, H). Figure 2G represents how a low spatial frequency pattern is selectively transmitted by a randomly distributed array of optical tubes. In this case individual tubes on the edge of a pattern that transmit either a white or black signal have a small effect on the total signal. However, in Figure 2H where a high spatial frequency grating is imaged the inherent noise in the system due to whether individual tubes at the edge of a pattern selectively transmit either a white or black signal has a very large effect on the overall signal. Thus noise in the distribution of the array of tubes contributes to limiting resolution of the system. The second factor that affects the resolution with which a mechano-optical lens array can resolve spatial information is the number of individual tubes that may selectively transmit rays of light from a part of a subject depending upon the working distance between the subject matter and the optical device. For example, Figure 3 shows that if a point in space is placed at position (A) near the aperture of an individual tube then light reflected from that particular point can only effectively be transmitted by one tube. However, if a point in space is placed at a distance of approximately the length of the tubes then light reflected from that point can be transmitted by multiple tubes (Figure 3; position B). If we consider a stack of tubes in a two dimensional array then the number of optical units that transmit light from a given point in space increases as a polynomial series where N (Number tubes viewing point in space) = 3x2 +3x + 1, where x is distance of point from the mechano-optical device. It is important to note that bees do not see stereo images in the way that primates do (Srinivasan 1993) and in this study it is reasonable to consider each eye as a separate unit for modelling purposes. It should also be noted that the model we have used simulates the frontal foveal region of a bee eye where the interommatidial angle is close to zero. Therefore the formula previously mentioned very closely represents the visual process of the bee. However, as the interommotidial increased towards the edges of the eye the system becomes more dynamic and difficult to model and the formula serves only as an approximation. Thus with increasing working distance from the mechano-optical device there is a rapidly increasing number of units in the array that view the stimulus, and so the ability to resolve the difference between points diminishes. This can be seen in the resolution of the flower in Figure 2 A-F. This empirical and theoretical data explains why a mechano-optical device is best suited to viewing objects at relatively small working distances. This study illustrates the potential problem of using mechano-optical type devices for imaging because of the sharp decline in the mtf, and the two main factors limiting the optical principle. However, this type of device is possibly useful in special applications where highly miniaturised optics are desirable (and other factors like diffraction come into consideration) (Land 1997), or it is possible to use the optics in confined localities where subject working distances are very small. For example, the interesting artificial compound eye recently built by Jeong et al. (2006) might potentially be useful in medical analysis type devices where the optics placed on an endoscope might permit wide angle views of nearby subject matter not possible with conventional optics (Jeong et al. 2006). Thus it appears that in addition to the lens and pinhole optical principles that evolved in nature and are useful for photography (Wald 1950), the compound optics common in many invertebrates like bees are potentially another solution that may have important real world photographic applications. Acknowledgements This material is based on research sponsored by the US Air Force Research Laboratory under agreement AOARD grant #064038&074080 to AGD. The U.S. Government is authorised to reproduce and distribute reprints for Government purposes notwithstanding any copyright notation thereon. The views and conclusions contained herein are those of the authors and should not be interpreted as necessarily representing the official policies or endorsements, either expressed or implied, of the Air Force Research Laboratory or the U.S. Government. References Dyer, A.G., Williams, S.K. 2005. A mechano-optical lens array to photographically simulate insect vision. Imaging Science Journal 53:209-213. Jander. U., and Jander, R. 2002. Allometry and resolution of bee eyes (Apoidea). Arthropod Structure and Development, Volume 30:179-193 Jeong, K., Kim, J., Lee, L.P. 2006. Biologically inspired artificial compound eyes. Science. 312:557-561 Knowles, A., Dartnall, H.J.A. 1977. The Photobiology of Vision. London: Academic Press Land, M.F. 1997. The resolution of insect compound eyes. Israel Journal of Plant Science 45:79-91 Muntz, W.R.A. and Raj, U. 1984. On the visual systems of Nautilus pompilius. Journal of Experimental Biology 109:253-261 Renner, E. 1999. Pinhole Photography: Rediscovering a Historic Technique. Oxford, UK: Focal Press.Shannon, C.E. 1998. Communication in the Presence of Noise. Proceedings of the IEEE 86:447-457. Stroebel, L., Compton, J., Current, I., and Zakia, R., 1996. Photographic Materials and Processes. London: Focal Press Srinivasan, M.V., Lehrer, M. 1988. Spatial acuity of honeybee vision and its spectral properties. Journal Comparative Physiology in America 162:159-172 Srinivasan, M.V., 1993. How insects infer range from visual motion. In: Visual Motion and its Role in the Stabilization of Gaze, F.A. Miles and J. Wallman (eds), Elsevier, Amsterdam, 139-156. Tanida, J., Shogenji, R., Kitamura, Y., Yamada, K., Miyamoto, M. and Miyatake, S., 2003. Color imaging with an integrated compound imaging system. Opt Express 11: 2109-2117. Wald, G.W. 1950. Eye and camera. Scientific American. 183:32-41 Williams, S.K., Dyer, A.G. 2002. Seeing through straws. In Images from Science, An Exhibition of Scientific Photography:108-109. Rochester, N.Y.: Cary Graphic Arts Press, Rochester Institute of Technology. Williams, S.K., Dyer, A.G. 2007. A photographic simulation of insect vision. The Journal of Ophthalmic Photography 29:10-14.
Susanne Williams completed a BSc at Monash University, Australia in 1998 and then a B App Sci (Photography) at RMIT, Australia in 2001. Between 2003 and 2007 she worked as an ophthalmic and medical photographer at The Royal Victorian Eye and Ear Hospital, Melbourne, Australia. She is Currently working as a 3D medical photographer at The Royal Children’s Hospital, Melbourne, Australia. Contact her at: susanne.williams@rch.org.au David Reser, B.S., M.S., Ph.D, is a Research Officer in the Department of Physiology of Monash University in Melbourne, Australia. Prior to this posting, he was a Senior Scientist with Allergan, Inc., and a Research Fellow at the Georgetown University Institute for Cognitive and Computational Science. He obtained his B.S. in Biology from the University of Connecticut, and his M.S. and Ph.D in Neuroscience from the Albert Einstein College of Medicine. His primary research interest is in the physiological mechanism of sensory information processing, especially with respect to communication between brain areas. Adrian Dyer completed a B App Sci in Photography at RMIT (Australia) in 1988, and then a Ph.D at RMIT in 2000. Between 1988 and 2001 he worked as a scientific photographer at Monash University (Australia). In 2002 he was awarded an Alexander von Humboldt Fellowship for research on visual processing in insects (Germany). He then completed postdoctoral fellowships on insect vision Mainz University (Germany) and Cambridge University (UK). Currently he is a postdoctoral Fellow at Monash University investigating information processing in bee miniature brains.
|
Copyright
2008, The Journal of Biocommunication, All Rights Reserved
Table
of Contents for VOLUME 34, NUMBER 1